জিন থেকে জিনোম— ক্রমবর্ধমান জটিলতার দিকে যাত্রা
- প্রকাশ: ০২:১৪:২৬ অপরাহ্ন, বৃহস্পতিবার, ২৫ অগাস্ট ২০২২
- / ১০৯৪ বার পড়া হয়েছে
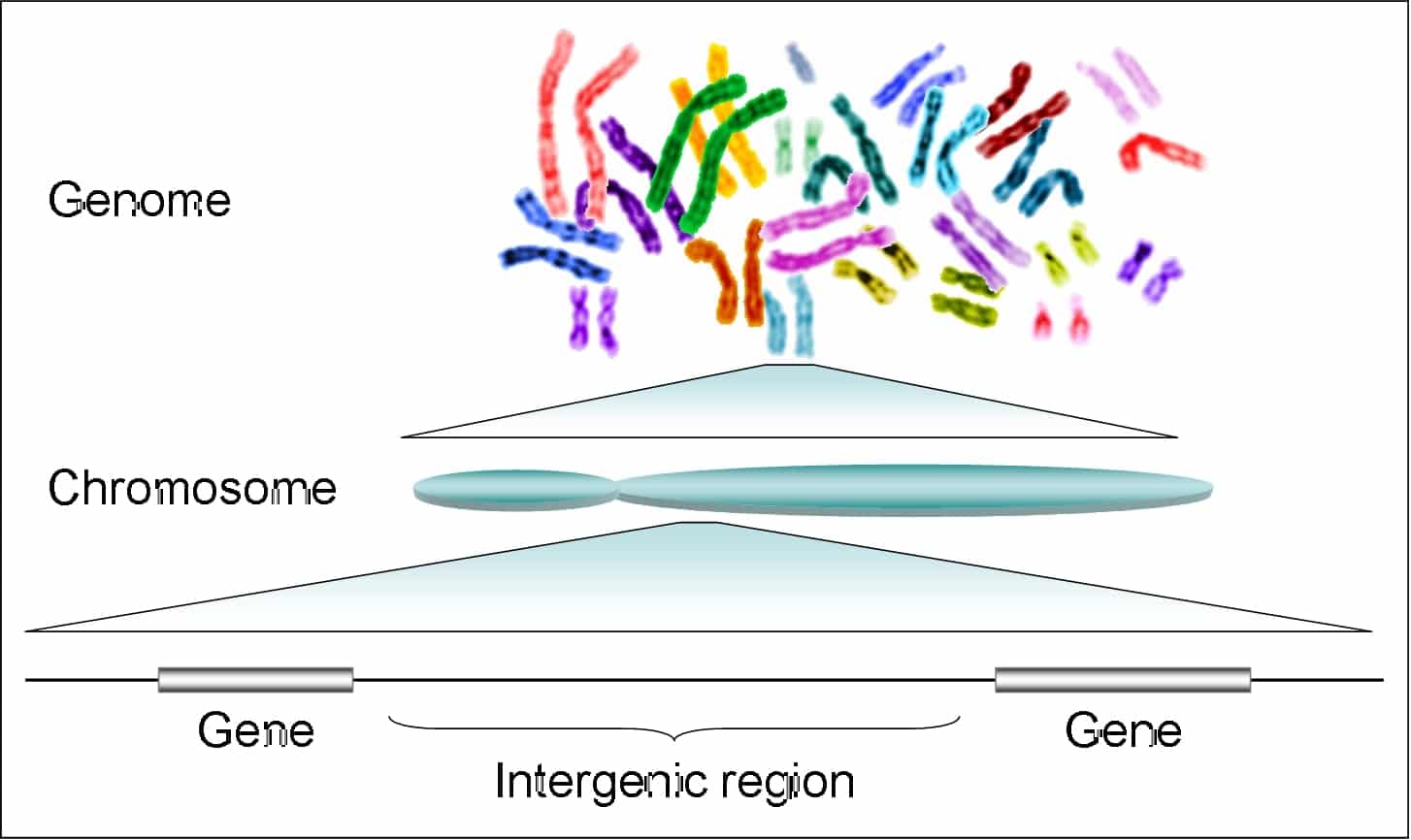
প্রতিটি জীবের (ব্যাকটেরিয়া, উদ্ভিদ ও প্রাণী) বংশাণুর বা বংশগতির মৌলিক আণবিক একক হচ্ছে জিন (gene), যা ক্রোমোজোমে অবস্থিত DNA/ডিএনএ (ডিঅক্সিরাইবো নিউক্লিক অ্যাসিড) অণুর একটি নির্দিষ্ট অংশ। জীবজগতের বংশগতির ধারক ও বাহক হচ্ছে ডিএনএ। জিন-এর মাধ্যমেই একটি জীবের বৈশিষ্ট্য নির্ধারিত হয়। একটি জিন হচ্ছে একটি গল্পের মত এবং ডিএনএ-র ক্রমগুলো হলো সেই ভাষা যে ভাষায় গল্পগুলো লেখা হয়েছে। সেই গল্পগুচ্ছে রয়েছে অসংখ্য প্রোটিনের কথা- জীবের বৃদ্ধি, বিপাক, আবেগপ্রবণতা, প্রজনন, পরিবেশের সাথে অভিযোজিত করা-সহ যাবতীয় প্রোটিন-নিয়ন্ত্রিত কার্যক্রমের কথা। মানবজিনোমের সম্পূর্ণ ডিএনএ-সিকোয়েন্সিং সম্পন্ন হয়েছে ২০২১ সালে। ডিএনএ-এর ক্রমবিন্যাসের তথ্য অনুযায়ী X-ক্রোমোজোমসহ হ্যাপ্লয়েড মানব জিনোম (২৩টি ক্রোমোজোম) প্রায় ৩ বিলিয়ন (৩,০৫৪,৮১৫,৪৭২ bp [বেস-পেয়ার]) নিউক্লিওটাইড নিয়ে গঠিত। এই নিখিল জিনোমে রয়েছে সর্বমোট ৬৩,৪৯৪টি জিন, যার মধ্যে মাত্র ১৯,৯৬৯টি জিন (জিনোমের মাত্র ১.৫%) হলো প্রোটিন-কোডিং (protein-coding) জিন, আর বাকি সবই নন-কোডিং RNA জিন। এসব জিনের প্রতিটিতে রয়েছে ভিন্ন ভিন্ন জৈব রাসায়নিক সংকেত, অর্থাৎ জীবের বংশগত বৈশিষ্ট্য যা বংশপরম্পরায় সঞ্চারিত হয়ে আসছে। তবে ক্রোমোজোমগুলোতে এইসব জিন একের পর এক পিঠাপিঠি গুছানো নয়, জিনগুলোর মধ্যবর্তী দূরত্ব বিস্তর। তাদের এই দূরত্ব পূরণ করেছে হাজার-হাজার নন-কোডিং নিউক্লিওটাইড, যা অনেকক্ষেত্রে অতিমাত্রায় পুনরাবৃত্তিমূলক (repetitive)। এদের বলা হয় স্পেসার ডিএনএ (spacer DNA) ও জিনোমে এদের সংখ্যা অগণিত।
এখানে পড়ুন
একটি জিনের গঠন ও জেনেটিক কোড
জিনোম বলতে আসলে একটি জীবের সামগ্রিক ডিএনএ-কে বোঝায়। একে সহজভাবে বলা চলে জীবনের নীলনকশা বা একটি জীবের জীবন-বিধান। একটি জিন কার্যতঃ কয়েকটি অঞ্চলে বিভক্ত: একটি নিয়ন্ত্রক অঞ্চল (যেমন, promoter) যা জিনের অভিব্যক্তিকে নিয়ন্ত্রণ করে, একটি প্রতিলিপি অঞ্চল (যার জেনেটিক কোডের সংকেত অনুযায়ী সংশ্লেষ বা প্রতিলিপিত হয় mRNAসহ অন্যান্য RNA) এবং নানান কার্যিক (regulatory) ক্রম অঞ্চল। আর, নোবেল বিজয়ী ফ্রান্সিস ক্রিকের সুপ্রতিষ্ঠিত সেই ‘সেন্ট্রাল ডগমা’ তো আমাদের সকলের-ই জানা, যা আণবিক জীববিজ্ঞানে আজও প্রামাণ্য তত্ত্ব বলে মান্যতা পায়- অর্থ্যাৎ, “একটি জিন থেকে তৈরি হয় এমআরএনএ (mRNA) এবং সেখান থেকে জেনেটিক কোডের তথ্য অনুযায়ী সংশ্লেষিত হয় একটি প্রোটিন” (হতে পারে একটি কাঠামোগত প্রোটিন, হরমোন বা এনজাইম) এবং এই ধারাটি অপ্রত্যাবর্তনীয়। তত্ত্বটি শোনায় যতটা সহজ, কার্যতঃ সেন্ট্রাল ডগমার দুটি স্তরই বেশ জটিল। বিবর্তনের ইতিহাসে সুকেন্দ্রিক (eukaryote) জীবে একটি জিনের প্রতিলিপি অঞ্চলে একাধিক প্রোটিন-কোডিং (coding) অঞ্চলের সাথে জড়ো হয়েছে নন-কোডিং (non-coding) অঞ্চল, যাদের বলা হয় যথাক্রমে একসন (Exon) ও ইন্ট্রোন (Intron), যদিও ভাইরাস (অ্যাডিনোভাইরাস) ও ব্যাকটেরিয়ার জিনগুলোতে কদাচিৎ ইন্ট্রোনের উপস্থিতি দেখা যায়। তবে ব্যাকটেরিয়ায় সেগুলো প্রোটিন-কোডিং অঞ্চলে উপস্থিত নয়। জিন বিশেষে দেখা গেছে এক্সন এবং ইন্ট্রোনের সংখ্যা ভিন্ন। উদাহরণস্বরূপঃ মানুষের হিস্টোন (Histone) 1A জিনের শুধুমাত্র একটি এক্সন (৭৮১ নিউক্লিওটাইড) আছে এবং সেখানে কোন ইন্ট্রন নেই, কিন্তু ডিস্ট্রোফিন (Dystrophin) জিনের ৭৯টি এক্সন জুড়ে রয়েছে ১০,৫০০ নিউক্লিওটাইড এবং ৭৮টি ইন্ট্রোনে রয়েছে ২,২০৯,৮৮১ নিউক্লিওটাইড। মানবজিনোমে সম্ভৱতঃ ডিস্ট্রোফিন-ই আকারে সবথেকে বড় জিন। প্রোটিন সংশ্লেষণে ইন্ট্রোনগুলোর কোনো সরাসরি ভুমিকা না থাকায় চূড়ান্ত এমআরএনএ (mRNA) অণুর পরিপক্কতার সময় ‘স্প্লাইসিং’ (RNA splicing) প্রক্রিয়া দ্বারা ইন্ট্রোনগুলোকে সরানো হয়। তবে গত বিশ বছরের গবেষণায় দেখা গেছে যে, প্রোটিন সংশ্লেষণে ইন্ট্রোনগুলোর কোনো সরাসরি ভুমিকা না থাকলেও, তারা জিনের অভিব্যক্তি নিয়ন্ত্রণের অবিচ্ছেদ্য অঙ্গ। বিভিন্ন ধরণের ক্যান্সার যথা স্তন, ডিম্বাশয় ও ফুসফুস ক্যান্সার সৃষ্টিতে ইন্ট্রোনের ভূমিকাও স্বীকৃত। বিংশ শতকে বংশাণু ও বংশগতি সম্পর্কে, বিশেষকরে ডিএনএর দ্বি-সূত্রক গঠন সম্পর্কে ধারণা আসার পর থেকে, জিন ও জিনোম নিয়ে আণবিক জীববিজ্ঞানীদের জানার পরিধি কল্পনাতীতভাবে প্রসার লাভ করেছে।

সর্বপ্রথম জেনেটিক কোড (একবচন ‘কোডন’)-র ধারণাটি দেন নোবেল বিজয়ী ফ্রান্সিস ক্রিক ও বিজ্ঞানী সিডনি ব্রেনার। ডিএনএ বা আরএনএ অণুর কাঠামোতে থাকে চার ধরনের নিউক্লিওটাইড বেস: অ্যাডেনিন, গুয়ানিন, সাইটোসিন ও থায়ামিন (আরএনএ অণুতে থায়ামিনের পরিবর্তে থাকে ইউরাসিল)। ডিএনএ বা আরএনএ অণুর তিনটি পরস্পর-সংলগ্ন নিউক্লিওটাইড একটি সাঙ্কেতিক একক বা কোডন গঠন করে, যা প্রোটিন সংশ্লেষণের পরবর্তী ধাপে কোন্ অ্যামিনো অ্যাসিড যোগ করতে হবে, তা নির্ধারণ করে। একটি এমআরএনএ (mRNA) প্রতিলিপি সর্বদা তৈরি হয় দুটি ডিএনএ স্ট্র্যান্ডের একটি থেকে, যাকে বলা হয় টেমপ্লেট স্ট্র্যান্ড (template strand)। অর্থাৎ, এমআরএনএ অণুটি ডিএনএ টেমপ্লেট স্ট্র্যান্ডের পরিপূরক। ডিএনএ-র দ্বিতীয় স্ট্র্যান্ড বা সূত্রকটিকে বলা হয় নন-টেমপ্লেট স্ট্র্যান্ড। প্রকৃতপক্ষে, এমআরএনএ কোডনগুলো-ই রাইবোজোম নামক অঙ্গাণুতে, নির্দিষ্ট ট্রান্সফার আরএনএ (tRNA) ও রাইবোজোমীয়-RNA (rRNA)র সহযোগিতায়, প্রোটিন সংশ্লেষণের সময় প্রোটিন-শৃঙ্খলের নির্দিষ্ট অ্যামিনো অ্যাসিড নির্ধারণ করে। একটি জেনেটিক কোডন সর্বদা তিন অক্ষর (নিউক্লিওটাইড) বিশিষ্ট বা ট্রিপলেট (triplet), যা শুধুমাত্র একটি অ্যামিনো এসিড নির্ধারণ করে। সকল জীবে নির্দিষ্ট একটি কোডন একই অ্যামিনো এসিড নির্ধারণ করে, যদিও প্লাস্টিড, মাইটোকন্ড্রিয়া, কিছু জীবাণু, এমনকি কিছু একাইনোডার্ম, ফিতাকৃমি ও সিলিয়া-যুক্ত প্রোটোজোয়া জীবও বেশ কিছু জেনেটিক কোড বিকল্পভাবে ব্যবহার করে, যা স্ট্যান্ডার্ড জেনেটিক কোডের থেকে আলাদা। যেমন, UAA, UAG এই স্টপকোডন দুটি এককোষী প্যারামেসিয়ামে সংকেত বহন করে গ্লুটামিন অ্যামিনো অ্যাসিডের জন্য। জিন বা বাহক আরএনএ (mRNA)তে সব মিলিয়ে ৬৪টি সম্ভাব্য কোডন রয়েছে (ছবি দেখুন)। এর মধ্যে ৬১টি কোডন প্রোটিন তৈরিতে ২০ ধরনের প্রোটিনোজেনিক (proteinogenic) অ্যামিনো অ্যাসিডের জন্য সংকেত বহন করে। অর্থাৎ, একটি অ্যামিনো অ্যাসিড দুই বা তার অধিক কোডন দ্বারা নির্দিষ্ট হতে পারে। উদাহরণস্বরূপঃ mRNA-তে GCU, GCC, GCA, GCG কোডনগুলো অ্যালানিন (alanine) নামক অ্যামিনো অ্যাসিডের জন্য সংকেত বহন করে। অ্যামিনো অ্যাসিড নির্ধারণী কোড ছাড়াও রয়েছে সুনির্দিষ্ট শুরু বা সূচনা কোডন (AUG) ও সমাপ্তি/স্টপ কোড (UAA, UAG ও UGA)। বাহক আরএনএ-র শেষোক্ত তিনটি কোডনের যে কোনো একটির উপস্থিতিতে প্রোটিন সংশ্লেষণ এর সমাপ্তি ঘটে। জেনেটিক কোডের সম্পূর্ণ অর্থ উদ্ধারের জন্য মার্শাল নিরেনবার্গ ও হরগোবিন্দ খোরানা ১৯৬৮ সালে নোবেল পুরস্কার পান। নোবেল পুরস্কারের আরও একজন অংশীদার ছিলেন রবার্ট উইলিয়াম হোলি, যিনি ডিএনএ এবং প্রোটিন সংশ্লেষণের সংযোগ স্থাপনকারী ট্রান্সফার আরএনএ (tRNA)র আণবিক গঠন বর্ণনা করেন।
জিন/আরএনএ স্প্লাইসিং (Gene/RNA Splicing)
পূর্বেই বলা হয়েছে, ডিএনএ অণুতে দুই ধরনের জিন বিদ্যমান: প্রোটিন-কোডিং (protein-coding) জিন ও নন-কোডিং জিন। উদাহরণস্বরূপঃ ব্যাকটেরিয়াতে, প্রোটিন-কোডিং অঞ্চলগুলো সাধারণত জিনোম (বংশাণুসমগ্র)-র ৮৮ শতাংশ জুড়ে রয়েছে। পক্ষান্তরে, মানবজিনোমে রয়েছে মাত্র ১.৫ থেকে ২% প্রোটিন-কোডিং অঞ্চল। অর্থাৎ, মানবজিনোমের প্রায় ৯৮ শতাংশ-ই হচ্ছে নন-কোডিং ডিএনএ (ncDNA), যার একটি অংশ (৬%) গঠন করেছে নন-কোডিং জিন এবং কিছু জিন নিয়ন্ত্রণকারী (regulatory) সিকোয়েন্স/অঞ্চল। নন-কোডিং ডিএনএ-অঞ্চলগুলো একটি জীবের ডিএনএ-অণুর অবিচ্ছেদ্য অংশ বটে, কিন্ত সেগুলো প্রোটিন সংশ্লেষণের জন্য কোনো সক্রিয় জেনেটিক কোড ধারণ করে না। তবে, কিছু নন-কোডিং জিন (৬%) কার্যকরী নন-কোডিং আরএনএ (ncRNA) অণুতে প্রতিলিপিত হয় (যেমন tRNA, rRNA এবং প্রাকৃতিক microRNA, PIWI/piRNA, si[short-interfering]RNA ও lnc[long non-coding]RNA, এবং আরও কিছু নিয়ন্ত্রক-আরএনএ)। একটি প্রোটিন-কোডিং জিনের দুই প্রান্তেও রয়েছে নন-কোডিং নিয়ন্ত্রক ক্রম, যেমন 5’UTR (UnTranslated Region) ও 3’UTR, যা জিনের অভিব্যক্তি নিয়ন্ত্রণ করে, কিন্তু প্রোটিন সংশ্লেষণে অংশ গ্রহণ করে না। এছাড়াও নন-কোডিং অঞ্চলে রয়েছে ডিএনএ অনুলিপনের উৎস (origin of replication), স্যাটেলাইট ডিএনএ (satellite DNA), ট্রান্সপোজন (transposon), স্যুডোজিন (pseudogene), সেন্ট্রোমেয়ার (centromere) এবং টেলোমেয়ার (telomere: ক্রোমোজোমের শেষপ্রান্তে পুনরাবৃত্তিমূলক ডিএনএ সিকোয়েন্স)। কোডিং ও নন-কোডিং অঞ্চল ছাড়া ডিএনএ-র বাকি যেসব অঞ্চল বা সিকোয়েন্স রয়েছে তার বেশিরভাগই অকার্যকর ক্রম বলে মনে করা হয়, যেমন ইন্ট্রোন (মানবজিনোমের ২৬%), স্যুডোজিন (মানবজিনোমে রয়েছে প্রায় ১৫,০০০) এবং আন্তঃজেনিক (intergenic) ডিএনএ ক্রম। তবে প্রমাণ মিলেছে যে এদের কিছুটা অংশ জিন নিয়ন্ত্রণে ভূমিকা রাখে। এছাড়াও, বিভিন্ন সময়ে মানবজিনোমে প্রবেশ করেছে নানান ভাইরাসের (বিশেষকরে, রেট্রোভাইরাস) ডিএনএ-টুকরো যা জিনোমের প্রায় ৮ শতাংশ জুড়ে রয়েছে।

দৃশ্যতঃ এই অকার্যকর অঞ্চলগুলো মানবজিনোমসহ অনেক ইউক্যারিওট জিনোমেই পরজীবীর মত কালক্রমে বাস করছে। অনেক বিজ্ঞানী মনে করেন এগুলো সম্পূর্ণটাই জাঙ্ক (junk) বা স্বার্থপর ডিএনএ যা যুগ-যুগান্তর জিনোমে অহেতুক সংরক্ষিত। হয়তো এসব অঞ্চলগুলোর কাজ বিজ্ঞানীদের কাছে এখনও অজানা।
একটি জিনের প্রোটিন-কোডিং অঞ্চলের যে অংশে সক্রিয় জেনেটিক কোড উপস্থিত সেগুলো-ই হলো এক্সন (Exon)। মানবজিনোমের প্রায় দুই শতাংশ জুড়ে রয়েছে এক্সন এবং তাদের ফাঁকে ফাঁকে রয়েছে ইন্ট্রোন (২৬%), যেখানে সক্রিয় জেনেটিক কোড ধারণ করে না। ইন্ট্রোনখন্ডগুলো হচ্ছে অপ্রয়োজনীয়, প্রোটিন সংশ্লেষণে তারা অংশগ্রহণ করে না। তাই, প্রোটিন সংশ্লেষণের পূর্বেই তাদের ছেঁটে ফেলতে হয়। অকার্যকর এসব ইন্ট্রোনখণ্ডগুলো কে সরিয়ে এক্সনগুলোকে জোড়া দিয়ে একটি কার্যকর পরিপক্ক (mature) mRNA ক্রমের পুনর্গঠনের প্রক্রিয়া-ই হচ্ছে ‘স্প্লাইসিং’ এবং সেটি ঘটে কোষের নিউক্লিয়াসে ও ট্রান্সক্রিপশন-পরবর্তী (post-transcriptional) সময়ে। ব্যাকটেরিয়ায় ক্ষুদ্র আকারের ডিএনএ থাকার কারণে তার ডিএনএ-র পুরো দৈর্ঘ্য জুড়ে কোডিং জিন ধারণ করে, সেখানে জিন বা mRNA স্প্লাইসিং হয় না। তবে, প্রোক্যারিওটিক জীবে স্প্লাইসিং ঘটে tRNA এবং অন্যান্য নন-কোডিং আরএনএ-গুলোতে। বর্তমানে এই ধারণা প্রতিষ্ঠিত যে, জিন স্প্লাইসিং কৌশল ব্যবহার করে একই জিন থেকে একাধিক বিভিন্ন প্রোটিন তৈরি করা সম্ভব। সে কারণেই মানবজিনোমের সক্ষমতা রয়েছে এক লক্ষেরও বেশি প্রোটিন তৈরি করবার, হয়তো সে নিমিত্তেই জিনোমে প্রোটিন-কোডিং জিন রয়েছে মাত্র ২২ হাজার এবং রয়েছে ইন্ট্রোনেরও প্রয়োজনীয়তা।
সংক্ষেপে, ডিএনএ-তে অবস্থিত জিনগুলো আরএনএ পলিমারেজ (RNA polymerase)-র মাধ্যমে প্রতিলিপন (ট্রান্সক্রিপশন) করে একটি প্রাথমিক মেসেঞ্জার RNA (pre-mRNA)। প্রাথমিক ট্রান্সক্রিপ্টটি একটি পরিপক্ক (mature) এমআরএনএ গঠনের জন্য ট্রান্সক্রিপশন-পরবর্তী কিছু পরিবর্তন আনে, যেমন mRNA-র দুই প্রান্তে প্রতিরক্ষামূলক সিকোয়েন্স তৈরি করা (5’-capping [7-methylguanosine, m7G] ও 3’-polyA tail), অপ্রয়োজনীয় নন-প্রোটিন কোডিং অঞ্চল ইন্ট্রোনগুলোর অপসারণ ইত্যাদি। ইন্ট্রোন অপসারণের পর অবশিষ্ট এক্সনগুলোকে একটি একক অবিচ্ছিন্ন mRNA টেমপ্লেট হিসেবে পুনরায় গঠন করতে সংযুক্ত করা হয়। এইসব পরিবর্তন, সংশোধন হবার পরেই পরিপক্ক mRNAটি রাইবোজোমের নিকট গৃহিত হয়, প্রোটিন সংশ্লেষণের জন্য। স্প্লাইসিং প্রতিক্রিয়াটি অনুঘটিত হয় বিভিন্ন প্রোটিন এবং ছোট-ছোট নিউক্লীয় RNA-র (snRNAs: U1-U6) সমন্বয়ে গঠিত রাইবোনিউক্লিওপ্রোটিন (ribonucleoprotein) কমপ্লেক্স বা স্প্লাইসিওজোম (spliceosome) দ্বারা। বিকল্প স্প্লাইসিং (alternative splicing)এর মাধ্যমে তৈরি হয় একটি জিন বা প্রাক-mRNA থেকে একাধিক ভিন্ন প্রোটিন। একটি স্বচ্ছ ও বিশুদ্ধ mRNA তৈরি করতে এতসব জটিল ধাপ অতিক্রম করতে হয়, খরচ হয় বেশ কিছু জৈবিক শক্তি। স্প্লাইসিং প্রতিক্রিয়াটির এসব জৈবিক ধাপগুলো আবিষ্কার করেছিলেন মার্কিন বিজ্ঞানী ফিলিপ শার্প এবং রিচার্ড জে. রবার্টস, ফলশ্রুতিতে এই গুণী বিজ্ঞানীদ্বয় নোবেল পুরস্কার পেয়েছিলেন ১৯৯৩ সালে।
জিনোম (genome)
আণবিক জীববিজ্ঞান এবং জেনেটিক্সের ভাষায় একটি ‘জিনোম’ বা ‘বংশাণুসমগ্র’ হলো একটি জীবের সমস্ত জিনগত বা বংশগতিমূলক তথ্য। অর্থাৎ কোনও জীবের বংশাণুসমগ্রে ওই জীবের বা প্রজাতির বংশগতিমূলক সমস্ত তথ্য সংরক্ষিত থাকে। প্রকৃতপক্ষে, জিনোম এর একটি সুনির্দিষ্ট সংজ্ঞা নিয়ে আসা খুবই কঠিন। জিনোম সাধারণত নিউক্লীয় ডিএনএ অণুগুলোকে বোঝায় যেগুলো একটি জীবের জিনগত তথ্য বহন করে। ব্যাকটেরিয়াতে সাধারণত একটি বা দুটি বড় ক্রোমোজোমীয় ডিএনএ অণু ছাড়াও তাদের জিনগত উপাদান হিসেবে ছোট প্লাজমিড (plasmid) অণুও থাকে যা গুরুত্বপূর্ণ জেনেটিক তথ্য বহন করে। এছাড়া, প্রায় সমস্ত সুকেন্দ্রিক (ইউক্যারিওট) জীবে নিউক্লীয় ডিএনএ-র বাইরেও মাইটোকন্ড্রিয়া এবং শৈবাল ও উদ্ভিদের ক্লোরোপ্লাস্টে ডিএনএ অণু থাকে। অধিকন্তু, প্রায় সমস্ত সুকেন্দ্রিক জীব হলো ডিপ্লয়েড, কিন্তু জিনোমে অন্তর্ভুক্ত থাকে সেক্স ক্রোমোজোমসহ এক সেট হ্যাপ্লয়েড ক্রোমোজোমের তথ্য। তাই, মানবজিনোম বলতে বোঝায় X ও Y উভয় ক্রোমোজোমসহ ২২টি অটোজোমের বংশানুসমগ্র। একটি ভাইরাসের জিনোমও হতে পারে ডিএনএ বা আরএনএ। একটি ক্রোমোজোম হলো একটি দীর্ঘ ডিএনএ পলিমার অণু, যাকে ঘিরে থাকে হিস্টোন প্রোটিনসহ অন্যান্য ক্রোমোজোমীয় প্রোটিন, যা ক্রোমাটিন নামক একটি ঘনীভূত কমপ্লেক্স গঠন করে। ডিএনএর সাংগঠনিক একক তথা মনোমার অণুগুলোকে নিউক্লিওটাইড বলে। মোট চার ধরণের নিউক্লিওটাইড (অ্যাডেনিন, থায়ামিন,গুয়ানিন ও সাইটোসিন) নিয়ে ডিএনএ গঠিত। একটি ডিএনএ অণুর ভেতর মনোমার নিউক্লিওটাইডগুলো নির্দিষ্ট অনুক্রমে সজ্জিত থাকে এবং সঙ্কেত প্রদান করে বংশগতিমূলক তথ্য, তৈরি হয় এনজাইম ও হরমোনসহ নানান জাতীয় প্রোটিন।
উল্লেখ্য, দেহের নানান প্রোটিন সংশ্লেষিত হয় যেসব জিনগুলোর নির্দেশনা পেয়ে তারা মানবজিনোমে প্রতিনিধিত্ব করে জিনোমের মাত্র ২ শতাংশ। আদিকোষীয় জীব যেমন ব্যাকটেরিয়া বা এককোষী ছত্রাক ঈস্ট (yeast)এ দেখেছি ৫/৬ হাজার প্রোটিন-কোডিং জিন, কিন্তু মানবজিনোমে দেখা যায় প্রায় ২০,০০০ (বিশ হাজার) প্রোটিন-কোডিং জিন)। তাহলে, জিনোমের দুর্বোধ্যতায় প্রোটিন-কোডিং জিনের সংখ্যার সাথে জিনোমের আকার সম্পর্কিত কিনা তা বিবেচনার আগে মানবজিনোমকে আণবিকভাবে ব্যবচ্ছেদ করে দেখা যেতে পারে সেখানে প্রোটিন-কোডিং জিন ছাড়া আর অন্যান্য কী কী অঞ্চল জিনোমকে করে তুলেছে রহস্যময় (ছবি দেখুন)।

ট্রান্সপোজেবল এলিমেন্ট (Transposable element, TE)
সমগ্র জিনোমজুড়ে রহস্যময় বেশ কিছু ডিএনএ-র খন্ডসমূহ ছুটোছুটি করে যেন চঞ্চলা হরিণীর মত। বিচরণের নিয়মনিষ্ঠতা নেই, এলোমেলোভাবে তারা তাদের স্থান পরিবর্তন করে। কোষে বা জিনোমে এদের বাধা দেবার মত কেউ নেই। এদের বলা হয় ট্রান্সপোজেবল এলিমেন্ট (TE)। এরা মানবজিনোমের অনেক বড় একটি অংশ, সমস্ত জিনোমের প্রায় অর্ধেক, ৪৫%। TE রয়েছে দুই ধরণের: ট্রান্সপোজন (transposon) ও রেট্রো-ট্রান্সপোজন (retro-transposon)। এই দুইয়ের মধ্যে পার্থক্য হলো ট্রান্সপোজন গুলো ‘কাট এন্ড পেস্ট’ (cut and pase) পদ্ধতি অনুসরণ করে স্থান বদল করে, জিনোমে এর সংখ্যা কম (জিনোমের ৩%)। গত শতাব্দীর চল্লিশের দশকে ভুট্টা নিয়ে গবেষণা করতে গিয়ে বারবারা ম্যাকক্লিনটক (Barbara McClintock) ডিএনএ-খন্ডগুলোর এই চলিষ্ণু বৈশিষ্ট্য অবলোকন করেছিলেন এবং তাঁর আবিষ্কার সনাতন জিনতত্ত্বে এনেছিল এক বিপ্লব, যদিও প্রায় ত্রিশটি বছর তাঁর আবিষ্কারে কেও গুরুত্ব দেয় নি। অবশেষে, এই বিষয়ে একটি আন্তর্জাতিক স্বীকৃতি পেয়ে তিনি নোবেল পুরস্কার পান ১৯৮৩ সালে। তিনি দেখিয়েছিলেন, প্রতিটি ট্রান্সপোজন এক বিশেষ ধরণের এনজাইম ‘ট্রান্সপোজেজ’ (transposase) তৈরি করে, যা ‘কাট এবং পেস্ট’ প্রক্রিয়া দ্বারা ট্রান্সপোজন-খন্ডটিকে জিনোমের অন্য অংশে জোড়া দিতে সহায়তা করে। কেটে দেয়া ও জোড়া দেয়ার শূন্যস্থানগুলো পূরণ করে ডিএনএ পলিমেরেজ ও ডিএনএ লাইগেজ এনজাইম। আশ্চর্যের বিষয় হচ্ছে এদের জঙ্গমতার পেছনে কোনো সঠিক নিয়মনীতি নেই, রীতিমত জিনোমে এক ধরণের হঠকারিতা।
ট্রান্সপোজেবল এলিমেন্টের দ্বিতীয় গ্ৰুপটি হলো রেট্রো-ট্রান্সপোজন (retro-transposon), এরা আকারে বেশ বড়। তারা স্থান বদল করে না বটে, কিন্তু ‘কপি এন্ড পেস্ট’ (copy and paste)এর মাধ্যমে শুধু তাদের সংখ্যা বৃদ্বি করে এবং জিনোমের এখানে-সেখানে নিজেদের একীভূত করে নেয়। এভাবেই মানবসহ সুকেন্দ্রিক জীবের জিনোম হয়েছে ক্রমবর্ধমান ও দুর্বোধ্য। মানবজিনোমে (৩ বিলিয়ন নিউক্লিওটাইড) রেট্রো-ট্রান্সপোজনগুলো জুড়ে রয়েছে প্রায় ৪২ শতাংশ (ছবি দেখুন)। রেট্রো-ট্রান্সপোজন তিনভাগে বিভক্ত: LINE (long interspersed nuclear elements, ≈৬ kb), SINE (Short interspersed nuclear elements, ≈৫০০ bp) ও LTR (long terminal repeats)-রেট্রো-ট্রান্সপোজন। LINE ও SINE গ্রুপে LTR থাকে না। রেট্রোভাইরাসের (যেমন, HIV) মতো LTR-রেট্রো-ট্রান্সপোজোন ও LINE এলিমেন্টগুলোতে ‘রিভার্স ট্রান্সক্রিপটেজ’ (reverse transcriptase) এনজাইম তৈরি করবার জন্য নিজস্ব জিন রয়েছে। তাই খুব সহজেই তাদের আরএনএ প্রতিলিপিগুলো (RNA transcripts) ডিএনএ-তে রূপান্তরিত হয়ে মূল জিনোমে প্রবেশ করে। ক্রমান্বয়ে, এভাবেই জিনোমের অবারিত প্রসার ঘটেছে, জিনোমে বেড়েছে নিউক্লিওটাইডের সংখ্যা, যার অধিকাংশই অকার্যকর বলে বিজ্ঞানীদের ধারণা। বর্তমানে, মানবজিনোমে LINE এলিমেন্ট রয়েছে প্রায় ৮ লক্ষ ৫০ হাজার কপি। SINE রেট্রোট্রান্সপোজন ‘রিভার্স ট্রান্সক্রিপটেজ’ তৈরির জিন ধারণ করে না, তাই তারা সহযোগিতা নেয় LINE থেকে সৃষ্ট ‘‘রিভার্স ট্রান্সক্রিপটেজ’ এনজাইমের। মানবজিনোমসহ সকল ইউক্যারিওট জিনোমে এই বিশাল একটি TE পরিবার মানবজিনোমে রয়েছে যুগ-যুগান্তর পরজীবী ডিএনএ-অনুক্রম হিসেবে। যেহেতু এরা উল্কার মত সঞ্চরণশীল, ছুটোছুটি করে নিয়ন্ত্রনহীন, তাই গুরুত্বপূর্ণ জিনগুলোর মধ্যে এদের অনুপ্রবেশ ভয়ানক রকমের রোগ (যেমন ক্যান্সার, নিউরো-ডিজেনেরেটিভ বা স্নায়বিক অবক্ষয়মূলক রোগ) ও জিনোম অস্থিরতার সৃষ্টি করে। প্রসঙ্গতঃ মানবজিনোমের নিজস্ব ডিএনএ কোনো ধরণের ‘রিভার্স ট্রান্সক্রিপটেজ’ জিন ধারণ করে না, এটি একটি রেট্রোভাইরাসের বৈশিষ্ট্য।
স্যাটেলাইট ডিএনএ (Satellite DNA)
নন-কোডিং ডিএনএ-র আরও একটি বড় অংশ (মানবজিনোমের ১০ থেকে ১৫%) হচ্ছে স্যাটেলাইট ডিএনএ। প্রোটিন তৈরির জন্য স্যাটেলাইট ডিএনএ কোনো ধরণের সংকেত বহন করে না, অর্থাৎ এই অঞ্চলেটি কোনো জেনেটিক কোড ধারণ করে না। এরা এক বা একাধিক নিউক্লিওটাইডের পুনরাবৃত্তিমূলক ক্রম এবং পুনরাবৃত্তিগুলো ডিএনএ-তে সরাসরি একে অপরের সংলগ্ন (tandem repeat)। এদের বড় একটি অঞ্চল নিয়েই গঠিত সেন্ট্রোমেয়ার (centromere), যা কোষ বিভাজনের সময় অপত্য ক্রোমাটিডগুলো পৃথকীকরণের জন্য গুরুত্বপূর্ণ একটি উপ-অঞ্চল ও টেলোমেয়ার (telomere)। এছাড়াও, হেটেরোক্রোমাটিনের প্রধান কাঠামোগত উপাদান গঠন করে স্যাটেলাইট ডিএনএ। স্যাটেলাইট ডিএনএর পুনরাবৃত্তিমূলক ক্রমের সংখ্যা অনুযায়ী ভাগ করা হয়েছে মিনিস্যাটেলাইট (minisatellite) এবং মাইক্রোস্যাটেলাইট (microsatellite) অঞ্চলে। যখন ১০ থেকে ৬০টি নিউক্লিওটাইডের পুনরাবৃত্তি হয়, তখন একে মিনিস্যাটেলাইট বলা হয়। আর, পুনরাবৃত্তির সংখ্যা আরও কম (>১০ nt) যেখানে , তারা মাইক্রোস্যাটেলাইট হিসেবে পরিচিত। GC-সমৃদ্ধ মিনিস্যাটেলাইটগুলো পুরো জিনোমজুড় পাওয়া যায় যত্রতত্র। মানবজিনোমের প্রায় ৯০% মিনিস্যাটেলাইট ক্রোমোজোমের টেলোমেয়ার অঞ্চলে পাওয়া যায়। সেখানে একটি ট্যান্ডেম পুনরাবৃত্তিমূলক ক্রম হচ্ছে: TTAGGG TTAGGG TTAGGG। অপরদিকে, একটি মাইক্রোস্যাটেলাইট হলো এক থেকে দশটি নিউক্লিওটাইড পর্যন্ত দৈর্ঘ্যের পুনরাবৃত্তিমূলক ক্রম। উদাহরণস্বরূপ, ক্রম TATATATATA হলো একটি ডাইনিউক্লিওটাইড মাইক্রোস্যাটেলাইট ও GTCGTCGTCGTCGTC একটি ট্রাইনিউক্লিওটাইড মাইক্রোস্যাটেলাইট। মাইক্রোস্যাটেলাইটগুলো জিন নিয়ন্ত্রক অঞ্চল ও কোডিং অঞ্চলসহ মানবজিনোম জুড়ে রয়েছে যেখানে-সেখানে। উদাহরণস্বরূপ: মানবজিনোমে রয়েছে প্রায় এক লক্ষ ডাইনিউক্লিওটাইড মাইক্রোস্যাটেলাইট। আদিকোষীয় ও প্রকৃতকোষীয় জীবে ট্যান্ডেম রিপিটগুলো জিন নিয়ন্ত্রণ (প্রতিলিপিকরণ) ও কোষ বিভাজনে অংশগ্রহণসহ অন্যান্য গুরুত্বপূর্ণ ভূমিকা রাখে, তাদের মিউটেশনের কারণে কোলন ক্যান্সারসহ অন্যান্য রোগও দেখা দিতে পারে।
টেলোমেয়ার (Telomere)
জিনোমের টেলোমেয়ার অংশটি বিশেষভাবে আলোচনার দাবি রাখে, সে কারণেই এখানে সংক্ষেপে এটি ব্যাখ্যাত হলো। একটি টেলোমেয়ার হলো ক্রোমোজোমের প্রান্তে অবস্থিত বিশেষ কিছু প্রোটিন (TRF1, TRF2, TIN2, POT1, TPP1, RAP)-সমন্বিত পুনরাবৃত্তিমূলক (repetitive) নিউক্লিওটাইড ক্রম (যেমন, TTAGGG-পুনরাবৃত্তি)গুলোর একটি অঞ্চল। একটি টেলোমেয়ারে ১৫,০০০ থেকে ২০,০০০ নিউক্লিওটাইড (nt) থাকতে পারে (যেখানে বয়বৃদ্ধের ডিএনএ-তে দেখা গেছে মাত্র ১,৫০০ nt) ও সমগ্র জিনোম-জুড়ে আছে প্রায় ০.১ শতাংশ। টেলোমেয়ার একটি বিস্তৃত জেনেটিক বৈশিষ্ট্য যা সাধারণত ইউক্যারিওট জিনোমে পাওয়া যায়, প্রোক্যারিওটিক জিনোমে নয়- কারণ অধিকাংশ আদিকোষীয় ক্রোমোজোম রৈখিক (linear) নয়, গোলাকার। টেলোমেয়ারগুলো ক্রমাগত অবক্ষয় থেকে ক্রোমোজোম ডিএনএ-র প্রান্তীয় অঞ্চলগুলোকে রক্ষা করে, পক্ষান্তরে ক্রোমোজোমেরও প্রতিরক্ষা দেয়। দমন করে প্রান্তীয় রিকম্বিনেশন (recombination) ও নিশ্চিত করে ক্রোমোজোমের অখণ্ডতা। সত্তর দশকের গোড়ার দিকে, রুশ বিজ্ঞানী আলেক্সি ওলোভনিকভ প্রথম আভাস দিয়েছিলেন যে, ক্রোমোজোমগুলোর প্রান্ত সম্পূর্ণরূপে অনুলিপন (replication) করতে পারে না। ওলোভনিকভ আরও জানালেন যে যতবার একটি কোষের বিভাজন হয়, ততবার-ই টেলোমেয়ারের প্রান্তীয় কিছু (ডিএনএ) নিউক্লিওটাইড হারিয়ে যায় এবং এক পর্যায়ে টেলোমেয়ার এত ছোট হয়ে যায় যখন আর কোষ বিভাজিত হয় না, অ্যাপোপটোসিস (apoptosis)র মাধ্যমে কোষগুলোর মৃত্যু হয়। অর্থাৎ, বার্ধক্যের সাথে সাথে টেলোমেয়ার ক্রমাগত ছোট হতে থাকে। ঘটনাটি বিস্ময়কর, তাই বিজ্ঞানীদের ভাবনার কারণ হয়ে দাঁড়িয়েছে। গবেষণায় দেখা গেছে, ধূমপান, মদ্যপান, অনিদ্রা, গোমাংস ভক্ষণ, স্থুলতা, অলস জীবন, ইত্যাদি টেলোমেয়ার ছোট হয়ে যাবার অন্যতম কারণ। দুৰ্ভাগ্যক্রমে, ওলোভনিকভ নোবেল পুরস্কার পান নি, নোবেলে ভূষিত হয়েছিলেন এলিজাবেথ ব্ল্যাকবার্ন (সে সময়ে ‘পোস্টডক্টরাল ফেলো’), ক্যারল গ্রেইডার ও জ্যাক সজোস্টাক ২০০৯ সালে। তাদের বিস্তর গবেষণা ছিল কীভাবে টেলোমেয়ার ও এনজাইম টেলোমেরেজ (telomerase) ক্রোমোজোমগুলোকে রক্ষা করে। কতিপয় কোষে (যেমন জননকোষ, স্টেম সেল ও ক্যান্সার কোষ) টেলোমেয়ার ছোট হতে দেখা যায় না এবং সেগুলোতে দেহকোষের তুলনায় টেলোমেরেজ এনজাইমের ঘনত্বও থাকে বেশি। ক্যান্সার নিয়ন্ত্রণ ও চিকিৎসায় বিজ্ঞানীরা টেলোমেয়ার ও টেলোমেরেজ এনজাইমকে কীভাবে দমন করা যায় (পক্ষান্তরে ডিএনএ অনুলিপন ও কোষ বিভাজন কীভাবে দমন করা যায়) তার সর্বোত্তম উপায় খোঁজার জন্য কঠোর পরিশ্রম করছেন।
জিনোমে স্যুডোজিনের পরিব্যাপ্তি
জিনোমের নন-কোডিং অঞ্চলের একটি নিষ্ক্রিয় গ্রুপ হচ্ছে স্যুডোজিন (Pseudogenes)। মুক্ত ব্যাকটেরিয়াগুলোর জিনোমে সাধারণতঃ স্যুডোজিন দেখা যায় না, তবে মিথোজীবী (symbionts) ও পরজীবী ব্যাক্টেরিয়াগুলোর জিনোমে বেশ কিছু স্যুডোজিন দেখা গিয়েছে। প্রকৃতকোষীয় জীবে (eukaryotes), বিশেষকরে স্তন্যপায়ী প্রাণীদের মধ্যে স্যুডোজিনের বড় একটি অংশ সমগ্র জিনোমজুড়ে অবস্থান করছে। বস্তুতঃ স্যুডোজিনগুলো প্রকৃত প্রোটিন-কোডিং জিনের নিষ্ক্রিয় কপি, প্রায়শই জিনের অনুলিপি (duplicate gene) দ্বারা উৎপন্ন, যা ফ্রেমশিফ্ট (frameshifts) মিউটেশন সঞ্চয়নের মাধ্যমে অকার্যকর হয়ে পড়েছে। এই গ্রুপের স্যুডোজিনগুলো সৃষ্টি হয়েছে জিন ডুপ্লিকেশন (gene duplication)র মাধ্যমে। এদের অবস্থান মাতৃজিনের পাশেও হতে পারে, অথবা একেবারেই ভিন্ন ক্রোমোজোমে। অধিকন্তু, ডুপ্লিকেশন (duplication) ছাড়াও স্বয়ং প্রোটিন-কোডিং জিনগুলোও সাধারণত স্টপ কোডন ও অন্যান্য কোডনে মিউটেশনের মাধমে নিষ্ক্রিয় হয়ে স্যুডোজিনে রূপান্তরিত হতে পারে। এদের বলা হয় একক (unitary) বা স্বতন্ত্র স্যুডোজিন। এই রকম একটি উদাহরণ হচ্ছে মানবজিনোমের ৮নং ক্রোমোজোমে স্থিত GULO (L-gulono-γ-lactone oxidase)জিন, যার নিষ্ক্রিয়তায় মানবদেহে ভিটামিন সি (vitamin C) আর সংশ্লেষ হতে পারে না। অধিকাংশ স্যুডোজিন mRNA-র জেনেটিক কোড অকার্যকর, ফলে সেগুলো প্রোটিনে অনুবাদিত হয় না, যদিও এর দু-একটি ব্যাতিক্রম রয়েছে, যেমন PTEN-P (tumor suppressor gene) স্যুডোজিন। মানবজিনোমে ‘ডুপ্লিকেট’ স্যুডোজিনের সংখ্যা আঠারো থেকে বিশ হাজার। অর্থ্যাৎ, জিনোমে কার্যক্ষম প্রোটিন-কোডিং জিন ও স্যুডোজিনের সংখ্যা প্রায় সমতুল্য। ডুপ্লিকেট ও স্বতন্ত্র (unitary) স্যুডোজিন ছাড়াও জিনোমে রয়েছে আরও একটি বড় শ্রেণীর স্যুডোজিন, যাদের বলা হয় প্রসেস্ড (processed) স্যুডোজিন। অর্থাৎ, প্রোটিন-কোডিং জিনের পরিপক্ক mRNA রিভার্স ট্রান্সক্রিপ্টেজ (reverse transcriptase) এনজাইম দ্বারা পুনরায় ডিএনএ-তে রূপান্তরিত হয় ও প্রবেশ করে মূল জিনোমে, পরিণত হয় স্যুডোজিনে, কারণ এদের জিনে কোনো প্রমোটর (promoter) অঞ্চল থাকে না। এই গ্রুপটি মূলত LINE রেট্রো-ট্রান্সপোজনকেই প্রতিনিধিত্ব করে (দেখুন ট্রান্সপোজেবল এলিমেন্ট)। মানবজিনোমে স্থায়ীভাবে অবস্থান করছে আট থেকে নয় হাজার প্রসেস্ড স্যুডোজিন। মানবজিনোমে স্যুডোজিনগুলো বিভিন্ন ক্রোমোজোমে ছড়ানো- কোনো ক্রোমোজোমে কম, কোথাও বেশি। যেমন, ১১নং ক্রোমোজোম-জুড়ে রয়েছে ৫৬% স্যুডোজিন। স্যুডোজিনের উদাহরণ দিতে গিয়ে একটি জিনপরিবারের কথা না বল্লেই নয়। মানবজিনোমে, ঘ্রাণজ (olfactory) রিসেপ্টর জিনপরিবারে রয়েছে ≈১০০০ জিন, যার মধ্যে ≈৬০০টি জিন-ই হচ্ছে অকার্যকর বা স্যুডোজিন। পক্ষান্তরে, ইঁদুর ও কুকুর প্রাণীদের ঘ্রাণজ রিসেপ্টর জিন পরিবারের স্যুডোজিন রয়েছে যথাক্রমে মাত্র ২০% ও ১৮%। বিজ্ঞানীদের গবেষণায় দেখা গেছে, প্রাইমেট (primates) প্রাণীদেরও রয়েছে আনুপাতিকভাবে কম স্যুডোজিন। তাই ওইসব প্রাণীদের ঘ্রাণশক্তি মানুষের ঘ্রাণশক্তির তুলনায় অনেক প্রখর। নন-কোডিং RNA জিনগুলোরও হয়তো অনেক স্যুডোজিন রয়েছে, কিন্তু জিনোমে তাদের শনাক্ত করা বেশ কঠিন। অধিকাংশ স্যুডোজিন অকার্যকর হলেও কতিপয় স্যুডোজিন স্বাভাবিকভাবে জিন নিয়ন্ত্রণে প্রয়োজনীয় ভূমিকা রাখে।
জিন-নিয়ন্ত্রণকারী ক্রম
একটি জিনের ৫’- ও ৩’- উভয় প্রান্তেই জিন-নিয়ন্ত্রণকারী ক্রম রয়েছে, যাদের মধ্যে ৫’-প্রান্তে অবস্থিত প্রোমোটর (promoter) অঞ্চলটি হচ্ছে অন্যতম, যা বিভিন্ন জিনে ১০০ থেকে ৪০,০০০ নিক্লিওটাইড দীর্ঘ হতে পারে। প্রোমোটর অঞ্চলটি প্রোটিন-কোডিং জিন এবং নন-প্রোটিন কোডিং জিনগুলোর অবিচ্ছেদ্য অংশ, যা যথাক্রমে mRNA এবং tRNA ও rRNA-গুলোর প্রতিলিপন (transcription)কে নিয়ন্ত্রণ করে। প্রসঙ্গত উল্লেখ্যঃ দ্বিসূত্রক ডিএনএ-র অ্যান্টিসেন্স স্ট্র্যান্ড (antisense strand)টি মেসেঞ্জার আরএনএ (mRNA) তৈরির টেমপ্লেট হিসেবে কাজ করে, যা একটি প্রোটিনের সংশ্লেষণকে নির্দেশ দেয়। মানবজিনোমে জিন-নিয়ন্ত্রণকারী ‘প্রোমোটর’ অঞ্চলের উল্লেখযোগ্য ও গুরুত্বপূর্ণ কিছু সিকোয়েন্স এখানে বিবৃত হয়েছে। কার্যতঃ প্রোমোটর এর তিনটি ভাগ রয়েছে: মূল (core) প্রোমোটর, নিকটবর্তী (proximal) প্রোমোটর ও দূরবর্তী (distal) প্রোমোটর। মূল প্রোমোটর অঞ্চলটির প্রধান ক্রম হলো ‘TATA box’ (‘টাটা’ বক্স)। প্রকৃতপক্ষে, এই বক্সটির কনসেন্সাস (consensus) সিকোয়েন্স হচ্ছে 5′-TATA(A/T)A(A/T)-3’। একটি জিনের ১৭bp-বিশিষ্ট ‘প্রতিলিপন প্রারম্ভিক সিকোয়েন্স’ (YYANWYY), যাকে বলা হয় ‘ইনিশিয়েটর এলিমেন্ট (initiator element, Inr), থেকে নিকটবর্তী অঞ্চলে, অর্থাৎ ২৫ থেকে ৩৫নিউক্লিওটাইড ডিএনএ-র সম্মুখদিক বা অগ্রভাগে (upstream), ‘টাটা বক্স’ অবস্থান করে। Inr সিকোয়েন্স দেখা যায় মানবজিনোমের ৪৯% জিন-প্রমোটরে। এই বক্সটিকে নন-কোডিং ডিএনএ সিকোয়েন্স হিসেবে বিবেচনা করা হয়, যা আর্কিয়া এবং ইউক্যারিওট জিনগুলোর মূল প্রোমোটর অঞ্চলের প্রারম্ভিক ভাগে পাওয়া ডিএনএর একটি গুরুত্বপূর্ণ ক্রম। তবে, অনেক জিনের প্রোমোটর অঞ্চলে ‘TATA বক্স’ থাকে না; গবেষণায় দেখা গেছে, মানবজিনোমে মাত্র ২৪% জিনের প্রোমোটর অঞ্চলে রয়েছে TATA বক্স। TATA বক্সের ব্যাকটেরিয়াল হোমোলগকে বলা হয় প্রিবনৌ (Pribnow)বক্স- ‘TATAAT’। একটি জিনের প্রতিলিপনের জন্য ‘TATA’ বক্সে যুক্ত হয় টাটা-বন্ধনকারী প্রোটিন (TATA-binding protein,TBP), TFIID ও নুন্যকল্পে আরও সাতটি ট্রান্সক্রিপশন ফ্যাক্টর। সাথে মিলিত হয় (প্রকৃতকোষীয় জিনগুলোতে) আরএনএ পলিমারেজ II (RNA polymerase II)। আর, RNA pol I ও RNA pol II প্রতিলিপন করে যথাক্রমে rRNA(ribosomal RNAs) ও tRNA (transfer RNA)। আদিকোষীয় জিনোমে প্রিবনৌ বক্সের সাথে যুক্ত হয় আরএনএ পলিমারেজ ও সিগ্মা ফ্যাক্টর। যেসব জিনে (যেমন, মানব IRF-1 [Interferon regulatory factor-1]) টাটা-বক্স থাকে না, সেখানে টাটা-এর পরিবর্তে DPE (downstream promoter element) সিকোয়েন্স থেকে শুরু হয় প্রতিলিপন। প্রতিলিপন প্রক্রিয়ার সমাপ্তি টানার জন্য একটি জিনের শেষপ্রান্তে (৩’) রয়েছে একটি টার্মিনেশন সিকোয়েন্স (TTTATT), যেখানে পৌঁছে RNA পলিমেরেজ এনজাইমের দায়িত্ব শেষ। নব্য mRNA অণুর ৩’-প্রান্তে যুক্ত হয় ≈ ২০০ nt বিশিষ্ট ‘poly A-tail’ এবং এই কাজটি সম্পন্ন করে টার্মিনেশন প্রোটিন কমপ্লেক্স (CPSF, CstF, PAP, PABII, CFI, CFII)। প্রোমোটর অঞ্চলে TATA বক্স ছাড়াও নিকটবর্তী ও দূরবর্তী অংশে রয়েছে বেশ কিছু খন্ড-খন্ড সিকোয়েন্স যাকে বলা হয় রিপ্রেসর (repressor) ও এনহ্যানসার (enhancer), যার মধ্যে একটি গুরুত্বপূর্ণ ক্রম হলো E-box (CACGTG), যেখানে যুক্ত হয় ট্রান্সক্রিপশন ফ্যাক্টর BMAL1-Clock এবং Myc। জৈবিক-ঘড়ি নিয়ন্ত্রিত জিনগুলোর প্রোমোটরে-ও E-box একটি কার্যকরী সিকোয়েন্স (Haque R, Ali FG, Biscoglia R, et al. J Neurochem, 2010, 113(5): 1296-306)। প্রমোটরে আরও একটি গুরুত্বপূর্ণ জিন-নিয়ন্ত্রণকারী সিকোয়েন্স হলো CpG আইল্যান্ড (islands), যা প্রায় ৭০% জিনের প্রোমোটরে পাওয়া যায়। যাহোক, জিনোমের প্রোমোটর ক্রমগুলো কখনও আরএনএ অণুতে প্রতিলিপিত হয় না, জিনের প্রকাশকে নিয়ন্ত্রণ করাই হচ্ছে এদের মুখ্য কাজ।
জিনোমের জিন-নিয়ন্ত্রণকারী প্রোমোটর ছাড়াও আরও একটি অঞ্চল রয়েছে, UTR বা Untranslated regions, যা প্রতিলিপিত হয় কিন্তু অনুবাদিত হয় না। অর্থাৎ, এরা প্রোটিন তৈরিতে সরাসরি অংশ গ্রহণ করে না। শুধুমাত্র মেসেঞ্জার আরএনএ (mRNA)-র দুই প্রান্তে এদের দেখা মিলে। এদের অবস্থান একটি জিনের বা mRNAর দুই প্রান্তে: 5′-UTR (লিডার সিকোয়েন্স) ও 3′-UTR (ট্রেলার সিকোয়েন্স)। একটি বাহক আরএনএ (mRNA) একটি জিন থেকে তথ্য নিয়ে যায় রাইবোজোমে, যেখানে তার জেনেটিক কোডে সজ্জিত তথ্যগুলো অনুবাদিত হয় প্রোটিনের অ্যামিনো অ্যাসিড শৃঙ্খলে। তবে, mRNA অণু থেকে UTR অঞ্চল দুটি, সাথে 5’-cap ও 3’-poly (A) tail -ও , অনুবাদিত হয় না, তাদের কাজ ভিন্ন। তবে, অনেকটা ‘সর্ষের ভিতর ভূত’ লুকিয়ে থাকার মত 5’UTR-এ একটি ব্যতিক্রম আছে। এখানে ব্যাকটেরিয়ার একটি বিশেষ বৈশিষ্ট্য লুকায়িত, যা মানবজিনোমে uORF (upstream open reading frame) নামে অভিহিত, যা স্বয়ং প্রোটিনে রূপান্তরিত হয়ে জিনোমের অন্যান্য ORF অঞ্চলগুলোর অভিব্যক্তিকে নিয়ন্ত্রণ করতে পারে। যেখানে আদিকোষীয় জিনোমে 5’-UTR ৫ থেকে ১০ নিউক্লিওটাইড দীর্ঘ, সেখানে প্রকৃতকোষীয় জিনোমে প্রজাতি ভেদে তার সংখ্যা কালক্রমে কয়েক হাজারে গিয়ে দাঁড়িয়েছে, তার ক’টা ক্রম-ই বা কাজের, তা নিয়ে বিজ্ঞানীদের উৎসুক মনে প্রশ্ন তো রয়েছেই। ব্যাক্টেরিয়ায় এই ধরণের 5’-UTR অঞ্চলকে বলা হয় ‘শাইন-ডেলগারনো সিকোয়েন্স (Shine–Dalgarno sequence, 5′-AGGAGGU-3′), যা রাইবোজোম বাঁধনের স্থান হিসেবেও পরিচিত। অনেকটা সমজাতীয় একটি ক্রম ‘কোজাক কনসেনসাস সিকোয়েন্স’ (Kozak consensus sequence, 5’-ACCAUGG-3’) রয়েছে ইউক্যারিয়ট mRNAতে, যার মধ্যেই রয়েছে প্রোটিন শুরুর সংকেত- স্টার্ট কোডন ‘AUG’, যা মিথিওনিন (methionine) অ্যামিনো এসিডকে কোড করার মাধ্যমে একটি প্রোটিন তৈরির যাত্রা শুরু হয়। এছাড়াও, মানবজিনোমের প্রায় ৩৫% জিনগুলোর ২/১টি ছোট-বড় ইন্ট্রোন (intron)ও অবস্থান করে 5’-UTR অঞ্চলে।
আর, 3’-UTR তুলনামূলকভাবে কিছুটা বড়, যা প্রায় ৬০০০ নিউক্লিওটাইড পর্যন্ত দীর্ঘ হতে পারে। প্রমাণ মিলেছে, অ-অনুবাদিত এই অঞ্চল দুইটি সম্পূর্ণভাবে অকার্যকর নয়। যেমন, 5’-UTR থেকে ইঙ্গিত পাওয়া যায় রাইবোজোম কোথা থেকে প্রোটিন সংশ্লেষ শুরু করবে, তেমনি মাইক্রো-RNA (microRNA/miRNA)জাতীয় ক্ষুদ্র আরএনএ-গুলো mRNA-র কার্জক্রমকেও নিয়ন্ত্রণ করে এই অঞ্চল থেকে। বিশেষকরে, 3’-UTR অঞ্চলটি এই ধরণের নিয়ন্ত্রণে অধিকতর নিবেদিত। 3′-UTR-এর নির্দিষ্ট স্থানগুলোতে আবদ্ধ হওয়ার মাধ্যমে, miRNA ও কিছু রিপ্রেসোর অণু প্রোটিন সংশ্লেষণে বাধা দিয়ে বা সরাসরি mRNA প্রতিলিপির অবক্ষয় ঘটিয়ে জিনের অভিব্যক্তি হ্রাস করতে পারে। এছাড়া, 3’-UTR-র শেষ প্রান্তে যুক্ত হয় প্রায় ২৫০ (AAAAAAA…..) নিউক্লিওটাইড -দীর্ঘ একটি poly (A) লেজ, যা mRNAকে কোষে সর্বব্যাপী নিউক্লিয়েজ এনজাইমগুলোর থেকে রক্ষা করে। অধিকন্তু, মানুষের mRNAগুলোর 5’- ও 3’-UTR অঞ্চলে SINE (অ্যালু এলিমেন্ট, Alu element) ও LINE-সহ যথাক্রমে ১২% ও ৩৬% পুনরাবৃত্তিমূলক সিকোয়েন্সও বিদ্যমান, যাদের প্রোটিন সংশ্লেষণে কোনো ভূমিকা আছে কিনা তা বিজ্ঞানীদের জানা নেই। 3’-UTR অঞ্চলটিতে মিউটেশনের কারণে ভয়ানক কিছু রোগের সৃষ্টি হতে পারে, যেমন থ্যাল্যাসেমিয়া, লিউকোমিয়া, কঞ্জেনিটাল হৃদরোগ ইত্যাদি।
উপসংহৃতি
এই প্রবন্ধে ব্যাখ্যা করা হয়েছে মানুষের শুধুমাত্র নিউক্লীয় জিনোম নিয়ে, যেখানে মাইটোকন্ড্রিয়ার জিনোম অন্তর্ভুক্ত নয়। তবুও জেনে রাখা প্রয়োজন যে মাইটোকন্ড্রিয়ার জিনোমে রয়েছে প্রায় ১৬,৫০০ নিউক্লিওটাইড। কোডিং ও নন-কোডিং অঞ্চলগুলোর মিশলেই গঠিত হয়েছে একটি জীবের জিনোম/বংশাণুসমগ্র। দেহের প্রতিটি কোষ একই বংশাণুসমগ্র বহন করে, তা যকৃতকোষ হোক কিংবা স্নায়ুকোষ; কিন্তু একটি নির্দিষ্ট টিস্যুর কোষগুলোতে শুধুমাত্র প্রয়োজনীয় বংশাণুগুলোই কার্যকর থাকে ও শুধুমাত্র টিস্যু-নির্দিষ্ট প্রোটিন তৈরি করে- আর বাকি জিনগুলো সেখানে থাকে নীরব। আদিকোষীয় জিনোমের জটিলতা প্রকৃতকোষীয় (ইউক্যারিওট) জীবের তুলনায় কম। তার মূল কারণ হলো আদিকোষীয় জিনোমগুলো মূলত অনন্য (অ-পুনরাবৃত্ত) ডিএনএ-সিকোয়েন্স দ্বারা গঠিত, যেখানে অন্তর্ভুক্ত রয়েছে জিনগুলোর একক-কপি বা অপেরন, যা mRNA, tRNA ও rRNA তৈরি করে। সেখানে নেই কোনো নন-কোডিং অঞ্চলের আতিশয্য। প্রকারান্তরে, মেরুদন্ডী, বিশেষ করে স্তন্যপায়ী জীবগুলোর জিনোমে কালানুক্রমে সঞ্চিত হয়েছে নানান ধরণের ডিএনএ-উপকরণ, যার অনেকটাই অকার্যকর বলে ধারণা বিজ্ঞানীদের। যেখানে বিভিন্ন ব্যাকটেরিয়ায় দেখা যায় ৫০০ থেকে ৫০০০ জিন, সেখানে স্তন্যপায়ী প্রাণীদের জিনের সংখ্যা স্বাভাবিকভাবেই অনেক বেশি- ১৮০০০ থেকে ৩০,০০০ (বর্তমান হিসেবে অনুযায়ী) এবং মানবজিনোমে প্রোটিন-কোডিং জিনের সংখ্যা প্রায় ২০ হাজার।
প্রকৃতকোষীয় (ইউক্যারিওট) জিনোমের ভাণ্ডারে বিশাল বৈচিত্র্য রয়েছে, তবে সে বিশালত্বে যে জটিলতাও অনেক তা সন্দেহাতীতভাবেই বলা যায়। জিনোমের আকার, প্রোটিন-কোডিং জিনের সংখ্যা, কিংবা অন্য কিছু এই দুর্বোধ্যতার কারণ কিনা, তা নির্ণয়ে বিজ্ঞানীরা বিভিন্ন পদ্ধতিতে বিশ্লেষণ করেছেন। বিশ্লেষণে দেখা গেছে, স্বয়ং জিনোমের আকার এবং জীবের জটিলতার মধ্যে যেমন পারস্পরিক সম্পর্কের উল্লেখযোগ্য অভাব রয়েছে, তেমন জিনোমের আকার ও জীবের প্রোটিন-কোডিং জিনের সংখ্যার পারস্পরিক সম্পর্কও অসামঞ্জস্যপূর্ণ (নিচের ছবিটি দেখুন)। অন্যথায়, মানুষের জিনোমে কমপক্ষে মার্বেল লাংফিশ (marbled lungfish, Protopterus aethiopicus) এর মতো ১৩২ বিলিয়ন নিউক্লিওটাইড (bp) আকারের ডিএনএ থাকতো কিংবা তারও বেশি, যেখানে লাংফিশে (lungfish) মাত্র ১২ থেকে ১৪ হাজার জিন বিদ্যমান। মানবজিনোমে রয়েছে মাত্র ৩ বিলিয়ন নিউক্লিওটাইড (bp)। অপরপক্ষে, ক্ষুদ্র পরজীবী ট্রাইকোমোনাস (Trichomonas vaginalis) এর মাত্র ১৬০ মিলিয়ন (বিপি) আকারের জিনোমে রয়েছে প্রায় ৬০,০০০ প্রোটিন-কোডিং জিন (যেকোনো ইউক্যারিওটিক জিনোমের ক্রমানুসারে সর্বাধিক সংখ্যক প্রোটিন-কোডিং জিন)। এই রকম অসামঞ্জস্যপূর্ণ উদাহরণ অনেক আছে। স্পষ্টতইঃ জিনোমের আকার বা প্রোটিন-কোডিং জিনের সংখ্যা কোনটাই একটি জীবের জিনোমিক বা জৈবিক জটিলতার সূচক নয়। বরং, বিশ্লেষণে দেখা গেছে জিনোমের এই জটিলতার মূল কারণে রয়েছে নন-কোডিং অঞ্চলগুলো – যার অনেকটাই সক্ষেপে ব্যাখ্যা করা হয়েছে। অনেকে জিনোমের এই জটিল অবস্থানের জন্য ফ্যাক্টর হিসেবে ইন্ট্রোন ও অন্যান্য নন-কোডিং RNA সিকোয়েন্সগুলোকে চিহ্নিত করেছেন। জিনোমে ইন্ট্রোনগুলোর হেরফের-ই (বিকল্প স্প্লাইসিং) ২০ হাজার জিন থেকে তৈরি হয় লক্ষাধিক প্রোটিন। সৃষ্টিকর্তাই জানেন এর রহস্য। অধিকন্তু, জিন-নিয়ন্ত্রণকারী অঞ্চলগুলো (যেমন এনহ্যান্সার, সাপ্রেসর ইত্যাদি) এত বৈচিত্রময় এবং অনাবিষ্কৃত যে সেগুলো নিয়েও প্রয়োজন অনেক গবেষণার।

শেয়ার করুন
লেখকতথ্য






২৮℅ ছাড় পেতে ৩০/০৬/২০২৪ তারিখের মধ্যে প্রোমো কোড “professional10” ব্যবহার করুন। বিস্তারিত জানতে ও ভর্তি হতে ক্লিক করুন এখানে।







